-
Ultrasound is increasingly being used to modulate the properties of
biological membranes for applications in drug delivery and neuromodulation.
While various studies have investigated the mechanical aspect of the
interaction such as acoustic absorption and membrane deformation, it is not
clear how these effects transduce into biological functions, for example,
changes in the permeability or the enzymatic activity of the membrane. A
critical aspect of the activity of an enzyme is the thermal fluctuations of its
solvation or hydration shell. Thermal fluctuations are also known to be
directly related to membrane permeability. Here solvation shell changes of
lipid membranes subject to an acoustic impulse were investigated using a
fluorescence probe, Laurdan. Laurdan was embedded in multi-lamellar lipid
vesicles in water, which were exposed to broadband pressure impulses of the
order of 1MPa peak amplitude and 10{\mu}s pulse duration. An instrument was
developed to monitor changes in the emission spectrum of the dye at two
wavelengths with sub-microsecond temporal resolution. The experiments show that
changes in the emission spectrum, and hence the fluctuations of the solvation
shell, are related to the changes in the thermodynamic state of the membrane
and correlated with the compression and rarefaction of the incident sound wave.
The results suggest that acoustic fields affect the state of a lipid membrane
and therefore can potentially modulate the kinetics of channels and proteins
embedded in the membrane.
-
Crosslinked semi-flexible and flexible filaments that are actively deformed
by molecular motors occur in various natural settings, such as the ordered
eukaryotic flagellum, and the disordered cytoskeleton. The deformation of these
composite systems is driven by active motor forces and resisted by passive
filament elasticity, and structural constraints due to permanent cross-links.
Using a mean field theory for a one-dimensional ordered system, we show that
the combination of motor activity and finite filament extensibility yields a
characteristic persistence length scale over which active strain decays. This
decay length is set by the ability of motors to respond to combination of the
weak extensional elasticity, passive shear resistance and the viscoelastic
properties of the motor assembly, and generalizes the notion of persistence in
purely thermal filaments to active systems.
-
A biological system consists of a variety of polymers that are synthesized
from monomers, by catalysis that exists only for some long polymers. It is
important to elucidate the emergence and sustenance of such autocatalytic
polymerization. We analyze here the stochastic polymerization reaction
dynamics, to investigate the transition time from a state with almost no
catalysts to a state with sufficient catalysts. We found an optimal volume that
minimizes this transition time, which agrees with the inverse of the catalyst
concentration at the unstable fixed point that separates the two states, as is
theoretically explained. Relevance to the origin of life is also discussed.
-
Measurement of the life time of attachments formed by a single microtubule
(MT) with a single kinetochore (kt) {\it in-vitro} under force-clamp conditions
had earlier revealed a catch-bond-like behavior. In the past the physical
origin of this apparently counter-intuitive phenomenon was traced to the nature
of the force-dependence of the (de-)polymerization kinetics of the MTs. Here
first the same model MT-kt attachment is subjected to external tension that
increases linearly with time until rupture occurs. In our {\it force-ramp}
experiments {\it in-silico}, the model displays the well known `mechanical
signatures' of a catch-bond probed by molecular force spectroscopy. Exploiting
this new evidence, we have further strengthened the analogy between MT-kt
attachments and common ligand-receptor bonds in spite of the crucial
differences in their underlying physical mechanisms. We then extend the
formalism to model the stochastic kinetics of an attachment formed by a bundle
of multiple parallel microtubules with a single kt considering the effect of
rebinding under force-clamp and force-ramp conditions. From numerical studies
of the model we predict the trends of variation of the mean life time and mean
rupture force with the increasing number of MTs in the bundle. Both the mean
life time and the mean rupture force display nontrivial nonlinear dependence on
the maximum number of MTs that can attach simultaneously to the same kt.
-
A new class of formal latent-variable stochastic processes called hidden
quantum models (HQM's) is defined in order to clarify the theoretical
foundations of ion channel signal processing. HQM's are based on quantum
stochastic processes which formalize time-dependent observation. They allow the
calculation of autocovariance functions which are essential for
frequency-domain signal processing. HQM's based on a particular type of
observation protocol called independent activated measurements are shown to to
be distributionally equivalent to hidden Markov models yet without an
underlying physical Markov process. Since the formal Markov processes are
non-physical, the theory of activated measurement allows merging energy-based
Eyring rate theories of ion channel behavior with the more common
phenomenological Markov kinetic schemes to form energy-modulated quantum
channels. Using the simplest quantum channel model consistent with neuronal
membrane voltage-clamp experiments, activation eigenenergies are calculated for
the Hodgkin-Huxley K+ and Na+ ion channels. It is also shown that maximizing
entropy under constrained activation energy yields noise spectral densities
approximating $S(f) \sim 1/f^\alpha$, thus offering a biophysical explanation
for the ubiquitous $1/f$-type in neurological signals.
-
The cytoskeleton protein actin assembles into large bundles when supporting
stresses in the cell, but grows into a fine branched network to induce cell
motion. Such self-organization processes are studied in artificial networks of
cytoskeleton proteins with thick actin bundles and large motor protein
aggregates to enable optical observation. The effect of motor aggregate size on
the cytoskeleton mechanical properties is studied here in networks comprised of
much smaller motor assemblies. Large motor protein clusters are known to
increase the stiffness of actin based networks by introducing tension and
additional cross-linking cites. We find that these effects are universal to
actin gels regardless of actin bundle thickness and motor aggregate size and
are relevant, therefore, to a wide range of cytoskeleton based cellular
processes. In contrast, motor induced active fluctuations depend significantly
on motor assembly size, featuring unique non-Gaussian statistics at high
concentrations of small assemblies.
-
Infection by many viruses begins with fusion of viral and cellular lipid
membranes, followed by entry of viral contents into the target cell and
ultimately, after many biochemical steps, integration of viral DNA into that of
the host cell. The early steps of membrane fusion and viral capsid entry are
mediated by adsorption to the cell surface, and receptor and coreceptor
binding. HIV-1 specifically targets CD4+ helper T-cells of the human immune
system and binds to the receptor CD4 and coreceptor CCR5 before fusion is
initiated. Previous experiments have been performed using a cell line
(293-Affinofile) in which the expression of CD4 and CCR5 concentration were
independently controlled. After exposure to HIV-1 of various strains, the
resulting infectivity was measured through the fraction of infected cells. To
design and evaluate the effectiveness of drug therapies that target the
inhibition of the entry processes, an accurate functional relationship between
the CD4/CCR5 concentrations and infectivity is desired in order to more
quantitatively analyze experimental data. We propose three kinetic models
describing the possible mechanistic processes involved in HIV entry and fit
their predictions to infectivity measurements, contrasting and comparing
different outcomes. Our approach allows interpretation of the clustering of
infectivity of different strains of HIV-1 in the space of mechanistic kinetic
parameters. Our model fitting also allows inference of nontrivial
stoichiometries of receptor and coreceptor binding and provides a framework
through which to quantitatively investigate the effectiveness of fusion
inhibitors and neutralizing antibodies.
-
The production of mechanical stresses in living organisms largely relies on
localized, force-generating active units embedded in filamentous matrices.
Numerical simulations of discrete fiber networks with fixed boundaries have
shown that buckling in the matrix dramatically amplifies the resulting active
stresses. Here we extend this result to a bucklable continuum elastic medium
subjected to an arbitrary external stress, and derive analytical expressions
for the active, nonlinear constitutive relations characterizing the full active
medium. Inserting these relations into popular "active gel" descriptions of
living tissues and the cytoskeleton will enable investigations into nonlinear
regimes previously inaccessible due to the phenomenological nature of these
theories.
-
Targeting the mitochondrial enzyme FoF1-ATP synthase and modulating its
catalytic activities with small molecules is a promising new approach for
treatment of autoimmune diseases. The immuno-modulatory compound Bz-423 is such
a drug that binds to subunit OSCP of the mitochondrial FoF1-ATP synthase and
induces apoptosis via increased reactive oxygen production in coupled, actively
respiring mitochondria. Here we review the experimental progress to reveal the
binding of Bz-423 to the mitochondrial target and discuss how subunit rotation
of FoF1-ATP synthase is affected by Bz-423. Briefly, we report how F\"orster
resonance energy transfer (FRET) can be employed to colocalize the enzyme and
the fluorescently tagged Bz-423 within the mitochondria of living cells with
nanometer resolution.
-
Telomeres, the nucleoprotein complexes at the termini of linear chromosomes,
are essential for the processes of end replication, end protection, and
chromatin segregation. The Mre11 complex is involved in multiple cellular roles
in DNA repair and structure in the regulation and function of telomere size
homeostasis. In this study, we characterize yeast telomere chromatin structure,
phenotypic heritability, and chromatin segregation in both wild-type [MRE11]
and A470 motif alleles. MRE11 strains confer a telomere size of 300 base pairs
of G+T irregular simple sequence repeats. This DNA and a portion of
subtelomeric DNA is embedded in a telosome: a MNase-resistant non-nucleosomal
particle. Chromatin immunoprecipitation shows a three to four-fold lower
occupancy of Mre11A470T proteins than wild-type proteins in telosomes.
Telosomes containing the Mre11A470T protein confer a greater resistance to
MNase digestion than wild-type telosomes. The integration of a wild-type MRE11
allele into an ectopic locus in the genome of an mre11A470T mutant and the
introduction of an mre11A470T allele at an ectopic site in a wild-type strain
lead to unexpectedly differing results. In each case, the replicated sister
chromatids inherit telosomes containing only the protein encoded by the genomic
mre11 locus, even in the presence of protein encoded by the opposing ectopic
allele. We hypothesize that the telosome segregates by a conservative
mechanism. These data support a mechanism for the linkage between sister
chromatid replication and maintenance of either identical mutant or identical
wild-type telosomes after replication of sister chromatids. These data suggest
the presence of an active mechanism for chromatin segregation in yeast.
-
Telomeres, the nucleoprotein complexes at the termini of linear chromosomes,
are essential for the processes of end replication, end-protection, and
chromatin segregation. The Mre11 complex is involved in multiple cellular roles
in DNA repair and structure in the regulation and function of telomere size
homeostasis. In this study, we characterize yeast telomere chromatin structure,
phenotypic heritability, and chromatin segregation in both wild-type [MRE11]
and A470 motif alleles. MRE11 strains confer a telomere size of 300 base pairs
of G+T irregular simple sequence repeats. This DNA and a portion of
subtelomeric DNA is embedded in a telosome: an MNase-resistant non-nucleosomal
particle. Chromatin immunoprecipitation shows a three to four-fold lower
occupancy of Mre11A470T proteins than wild-type proteins in telosomes.
Telosomes containing the Mre11A470T protein confer a greater resistance to
MNase digestion than wild-type telosomes. The integration of a wild-type MRE11
allele into an ectopic locus in the genome of a mre11A470T mutant and the
introduction of a mre11A470T allele at an ectopic site in a wild-type strain
lead to unexpectedly differing results. In each case, the replicated sister
chromatids inherit telosomes containing only the protein encoded by the genomic
mre11 locus, even in the presence of protein encoded by the opposing ectopic
allele. We hypothesize that the telosome segregates by a conservative
mechanism. These data support a mechanism for the linkage between sister
chromatid replication and maintenance of either identical mutant or identical
wild-type telosomes after replication of sister chromatids. These data suggest
the presence of an active mechanism for chromatin segregation in yeast.
-
Virus capsids in interchromatin corrals of a cell nucleus are experimentally
known to exhibit anomalous diffusion as well as normal diffusion, leading to
the Gaussian distribution of the diffusion-exponent fluctuations over the
corrals. Here, the sojourn-time distribution of the virus capsid in local areas
of the corral, i.e., probability distribution of the sojourn time
characterizing diffusion in the local areas, is examined. Such an area is
regarded as a virtual cubic block, the diffusion property in which is normal or
anomalous. The distribution, in which the Gaussian fluctuation is incorporated,
is shown to tend to slowly decay. Then, the block-size dependence of average
sojourn time is discussed. A comment is also made on (non-)Markovianity of the
process of moving through the blocks.
-
All living systems can function only far away from equilibrium, and for this
reason chemical kinetic methods are critically important for uncovering the
mechanisms of biological processes. Here we present a new theoretical method of
investigating dynamics of protein-DNA interactions, which govern all major
biological processes. It is based on a first-passage analysis of biochemical
and biophysical transitions, and it provides a fully analytic description of
the processes. Our approach is explained for the case of a single protein
searching for a specific binding site on DNA. In addition, the application of
the method to investigations of the effect of DNA sequence heterogeneity, and
the role multiple targets and traps in the protein search dynamics are
discussed.
-
Mutations in the MTM1 gene, encoding the phosphoinositide phosphatase
myotubularin, are responsible for the X-linked centronuclear myopathy (XLCNM)
or X-linked myotubular myopathy (XLMTM). The MTM1 gene was first identified in
1996 and its function as a PtdIns3P and PtdIns(,5)P2 phosphatase was discovered
in 2000. In recent years, very important progress has been made to set up good
models to study MTM1 and the XLCNM disease such as knockout or knockin mice,
the Labrador Retriever dog, the zebrafish and the yeast Saccharomyces
cerevisiae. These helped to better understand the cellular function of MTM1 and
of its four conserved domains: PH-GRAM (Pleckstrin
Homology-Glucosyltransferase, Rab-like GTPase Activator and Myotubularin), RID
(Rac1-Induced recruitment Domain), PTP/DSP (Protein Tyrosine
Phosphatase/Dual-Specificity Phosphatase) and SID (SET-protein Interaction
Domain). This review presents the cellular function of human myotubularin MTM1
and its yeast homolog yeast protein Ymr1, and the role of MTM1 in the
centronuclear myopathy (CNM) disease.
-
The yeast Saccharomyces cerevisiae is one of the best characterized
eukaryotic models. The secretory pathway was the first trafficking pathway
clearly understood mainly thanks to the work done in the laboratory of Randy
Schekman in the 1980s. They have isolated yeast sec mutants unable to secrete
an extracellular enzyme and these SEC genes were identified as encoding key
effectors of the secretory machinery. For this work, the 2013 Nobel Prize in
Physiology and Medicine has been awarded to Randy Schekman; the prize is shared
with James Rothman and Thomas S{\"u}dhof. Here, we present the different
trafficking pathways of yeast S. cerevisiae. At the Golgi apparatus newly
synthesized proteins are sorted between those transported to the plasma
membrane (PM), or the external medium, via the exocytosis or secretory pathway
(SEC), and those targeted to the vacuole either through endosomes (vacuolar
protein sorting or VPS pathway) or directly (alkaline phosphatase or ALP
pathway). Plasma membrane proteins can be internalized by endocytosis (END) and
transported to endosomes where they are sorted between those targeted for
vacuolar degradation and those redirected to the Golgi (recycling or RCY
pathway). Studies in yeast S. cerevisiae allowed the identification of most of
the known effectors, protein complexes, and trafficking pathways in eukaryotic
cells, and most of them are conserved among eukaryotes.
-
A nine state master equation for Na+ channel gating that describes the
coupling between three activation sensors, a two-stage fast inactivation
process and a slow inactivated state, may be reduced to a system of kinetic
equations for a six state system. Assuming the activation sensors are
independent and the activation variable m has a faster relaxation than the
inactivation variable h, the open state probability during the action potential
may be expressed as $m(t)^3 h(t)$, and the kinetic equations may be reduced to
rate equations for activation, and fast and slow inactivation that describe
spike frequency adaptation in the neural membrane, a repetitive bursting
oscillation and a cardiac action potential with a plateau oscillation.
-
It is shown that quantum sustainability is a universal phenomenon which
emerges during environment-assisted electronic excitation energy transfer (EET)
in photobiological complexes (PBCs), such as photosynthetic reaction centers
and centers of melanogenesis. We demonstrate that quantum photobiological
systems must be sustainable for them to simultaneously endure continuous energy
transfer and keep their internal structure from destruction or critical
instability. These quantum effects occur due to the interaction of PBCs with
their environment which can be described by means of the reduced density
operator and effective non-Hermitian Hamiltonian (NH). Sustainable NH models of
EET predict the coherence beats, followed by the decrease of coherence down to
a small, yet non-zero value. This indicates that in sustainable PBCs, quantum
effects survive on a much larger time scale than the energy relaxation of an
exciton. We show that sustainable evolution significantly lowers the entropy of
PBCs and improves the speed and capacity of EET.
-
We propose a stochastic model for gene transcription coupled to DNA
supercoiling, where we incorporate the experimental observation that
polymerases create supercoiling as they unwind the DNA helix, and that these
enzymes bind more favourably to regions where the genome is unwound. Within
this model, we show that when the transcriptionally induced flux of
supercoiling increases, there is a sharp crossover from a regime where
torsional stresses relax quickly and gene transcription is random, to one where
gene expression is highly correlated and tightly regulated by supercoiling. In
the latter regime, the model displays transcriptional bursts, waves of
supercoiling, and up-regulation of divergent or bidirectional genes. It also
predicts that topological enzymes which relax twist and writhe should provide a
pathway to down-regulate transcription. This article has been published in
Physical Review Letters, May 2016.
-
Living cells dynamically modulate the local morphologies of their actin
cytoskeletons to perform biological functions, including force transduction,
intracellular transport, and cell division. A major challenge is to understand
how diverse structures of the actin cytoskeleton are assembled from a limited
set of molecular building blocks. Here we study the spontaneous self-assembly
of a minimal model of cytoskeletal materials, consisting of semiflexible actin
filaments, crosslinkers, and molecular motors. Using coarse-grained
simulations, we demonstrate that by changing concentrations and kinetics of
crosslinkers and motors we can generate three distinct structural phases of
actomyosin assemblies: bundled, polarity-sorted, and contracted. We introduce
new metrics to distinguish these structural phases and demonstrate their
functional roles. We find that the binding kinetics of motors and crosslinkers
can be tuned to optimize contractile force generation, motor transport, and
mechanical response. By quantitatively characterizing the relationships between
modes of cytoskeletal self-assembly, the resulting structures, and their
functional consequences, our work suggests new principles for the design of
active materials.
-
The availability of protein is an important factor for the determination of
the size of the mitotic spindle. Involved in spindle-size regulation is
kinesin-8, a molecular motor and microtubule (MT) depolymerase, which is known
to tightly control MT length. Here, we propose and analyze a theoretical model
in which kinesin-induced MT depolymerization competes with spontaneous
polymerization while supplies of both tubulin and kinesin are limited. In
contrast to previous studies where resources were unconstrained, we find that,
for a wide range of concentrations, MT length regulation is bistable. We test
our predictions by conducting in vitro experiments, and find that the bistable
behavior manifests in a bimodal MT length distribution.
-
Inside cells, cargos such as vesicles and organelles are transported by
molecular motors to their correct locations via active motion on cytoskeletal
tracks and passive, Brownian diffusion. During the transportation of cargos,
motor-cargo complexes (MCC) navigate the confining and crowded environment of
the cytoskeletal network and other macromolecules. Motivated by this, we study
a minimal two-state model of motor-driven cargo transport in confinement and
predict transport properties that can be tested in experiments. We assume that
the motion of the MCC is directly affected by the entropic barrier due to
confinement if it is in the passive, unbound state, but not in the active,
bound state where it moves with a constant bound velocity. We construct a
lattice model based on a Fokker Planck description of the two-state system,
study it using a kinetic Monte Carlo method and compare our numerical results
with analytical expressions for a mean field limit. We find that the effect of
confinement strongly depends on the bound velocity and the binding kinetics of
the MCC. Confinement effectively reduces the effective diffusivity and average
velocity, except when it results in an enhanced average binding rate and
thereby leads to a larger average velocity than when unconfined.
-
The intracellular environment is a dynamic space filled with various
organelles moving in all directions. Included in this diverse group of
organelles are vesicles, which are involved in transport of molecular cargo
throughout the cell. Vesicles move in either a directed or non-directed
fashion, often depending on interactions with cytoskeletal proteins such as
microtubules, actin filaments, and molecular motors. How these proteins affect
the local fluctuations of vesicles in the cytoplasm is not clear since they
have the potential to both facilitate and impede movement. Here we show that
vesicle mobility is significantly affected by myosin-II, even though it is not
a cargo transport motor. We find that myosin-II activity increases the
effective diffusivity of vesicles and its inhibition facilitates longer states
of non-directed motion. Our study suggests that altering myosin-II activity in
the cytoplasm of cells can modulate the mobility of vesicles, providing a
possible mechanism for cells to dynamically tune the cytoplasmic environment in
space and time.
-
During the asymmetric division of the Caenorhabditis elegans nematode zygote,
the polarity cues distribution and daughter cell fates depend on the correct
positioning of the mitotic spindle, which results from both centering and
cortical pulling forces. Revealed by anaphase spindle rocking, these pulling
forces are regulated by the force generator dynamics, which are in turn
consequent of mitotic progression. We found a novel, additional, regulation of
these forces by the spindle position. It controls astral microtubule
availability at the cortex, on which the active force generators can pull.
Importantly, this positional control relies on the polarity dependent LET-99
cortical band, which restricts or concentrates generators to a posterior
crescent. We ascribed this control to the microtubule dynamics at the cortex.
Indeed, in mapping the cortical contacts, we found a correlation between the
centrosome-cortex distance and the microtubule contact density. In turn, it
modulates pulling force generator activity. We modelled this control,
predicting and experimentally validating that the posterior crescent extent
controlled where the anaphase oscillations started, in addition to mitotic
progression. Finally, we propose that spatially restricting force generator to
a posterior crescent sets the spindle's final position, reflecting polarity
through the LET-99 dependent restriction of force generators to a posterior
crescent. This regulation superimposes that of force generator processivity.
This novel control confers a low dependence on microtubule and active force
generator exact numbers or dynamics, provided that they exceed the threshold
needed for posterior displacement. Interestingly, this robustness originates in
cell mechanics rather than biochemical networks.
-
During the early developmental process of organisms, the formation of the
left-right laterality requires a subtle mechanism, as it is associated with
other principal body axes. Any inherent chiral feature in an egg cell can in
principal trigger this spontaneous breaking of chiral symmetry. Individual
microtubules, major cytoskeletal filaments, are known as chiral objects.
However, to date there lacks convincing evidence of a hierarchical connection
of the molecular nature of microtubules to large-scale chirality, particularly
at the length scale of an entire cell. Here we assemble an in-vitro active
layer, consisting of microtubules and kinesin motor proteins, on a glass
surface. Upon inclusion of methyl cellulose, the layered system exhibits a
long-range active nematic phase, characterized by the global alignment of
gliding MTs. This nematic order spans over the entire system size in the
millimeter range and, remarkably, allows hidden collective chirality to emerge
as counterclockwise global rotation of the active MT layer. The analysis based
on our theoretical model suggests that the emerging global nematic order
results from the local alignment of MTs, stabilized by methyl cellulose. It
also suggests that the global rotation arises from the MTs' intrinsic
curvature, leading to preferential handedness. Given its flexibility, this
layered in-vitro cytoskeletal system enables the study of membranous protein
behavior responsible for important cellular developmental processes.
-
Switching of the direction of flagella rotations is the key control mechanism
governing the chemotactic activity of E. coli and many other bacteria.
Power-law distributions of switching times are most peculiar because their
emergence cannot be deduced from simple thermodynamic arguments. Recently it
was suggested that by adding finite-time correlations into Gaussian
fluctuations regulating the energy height of barrier between the two rotation
states, one can generate a power-law switching statistics. By using a simple
model of a regulatory pathway, we demonstrate that the required amount of
correlated `noise' can be produced by finite number fluctuations of reacting
protein molecules, a condition common to the intracellular chemistry. The
corresponding power-law exponent appears as a tunable characteristic controlled
by parameters of the regulatory pathway network such as equilibrium number of
molecules, sensitivities, and the characteristic relaxation time.





 Crosslinked semi-flexible and flexible filaments that are actively deformed by molecular motors occur in various natural settings, such as the ordered eukaryotic flagellum, and the disordered cytoskeleton. The deformation of these composite systems is driven by active motor forces and resisted by passive filament elasticity, and structural constraints due to permanent cross-links. Using a mean field theory for a one-dimensional ordered system, we show that the combination of motor activity and finite filament extensibility yields a characteristic persistence length scale over which active strain decays. This decay length is set by the ability of motors to respond to combination of the weak extensional elasticity, passive shear resistance and the viscoelastic properties of the motor assembly, and generalizes the notion of persistence in purely thermal filaments to active systems.
Crosslinked semi-flexible and flexible filaments that are actively deformed by molecular motors occur in various natural settings, such as the ordered eukaryotic flagellum, and the disordered cytoskeleton. The deformation of these composite systems is driven by active motor forces and resisted by passive filament elasticity, and structural constraints due to permanent cross-links. Using a mean field theory for a one-dimensional ordered system, we show that the combination of motor activity and finite filament extensibility yields a characteristic persistence length scale over which active strain decays. This decay length is set by the ability of motors to respond to combination of the weak extensional elasticity, passive shear resistance and the viscoelastic properties of the motor assembly, and generalizes the notion of persistence in purely thermal filaments to active systems.

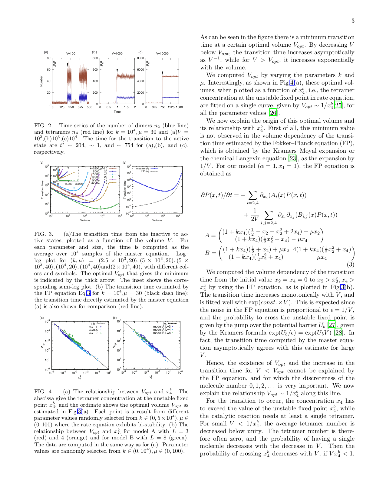


 A biological system consists of a variety of polymers that are synthesized from monomers, by catalysis that exists only for some long polymers. It is important to elucidate the emergence and sustenance of such autocatalytic polymerization. We analyze here the stochastic polymerization reaction dynamics, to investigate the transition time from a state with almost no catalysts to a state with sufficient catalysts. We found an optimal volume that minimizes this transition time, which agrees with the inverse of the catalyst concentration at the unstable fixed point that separates the two states, as is theoretically explained. Relevance to the origin of life is also discussed.
A biological system consists of a variety of polymers that are synthesized from monomers, by catalysis that exists only for some long polymers. It is important to elucidate the emergence and sustenance of such autocatalytic polymerization. We analyze here the stochastic polymerization reaction dynamics, to investigate the transition time from a state with almost no catalysts to a state with sufficient catalysts. We found an optimal volume that minimizes this transition time, which agrees with the inverse of the catalyst concentration at the unstable fixed point that separates the two states, as is theoretically explained. Relevance to the origin of life is also discussed.




 Measurement of the life time of attachments formed by a single microtubule (MT) with a single kinetochore (kt) {\it in-vitro} under force-clamp conditions had earlier revealed a catch-bond-like behavior. In the past the physical origin of this apparently counter-intuitive phenomenon was traced to the nature of the force-dependence of the (de-)polymerization kinetics of the MTs. Here first the same model MT-kt attachment is subjected to external tension that increases linearly with time until rupture occurs. In our {\it force-ramp} experiments {\it in-silico}, the model displays the well known `mechanical signatures' of a catch-bond probed by molecular force spectroscopy. Exploiting this new evidence, we have further strengthened the analogy between MT-kt attachments and common ligand-receptor bonds in spite of the crucial differences in their underlying physical mechanisms. We then extend the formalism to model the stochastic kinetics of an attachment formed by a bundle of multiple parallel microtubules with a single kt considering the effect of rebinding under force-clamp and force-ramp conditions. From numerical studies of the model we predict the trends of variation of the mean life time and mean rupture force with the increasing number of MTs in the bundle. Both the mean life time and the mean rupture force display nontrivial nonlinear dependence on the maximum number of MTs that can attach simultaneously to the same kt.
Measurement of the life time of attachments formed by a single microtubule (MT) with a single kinetochore (kt) {\it in-vitro} under force-clamp conditions had earlier revealed a catch-bond-like behavior. In the past the physical origin of this apparently counter-intuitive phenomenon was traced to the nature of the force-dependence of the (de-)polymerization kinetics of the MTs. Here first the same model MT-kt attachment is subjected to external tension that increases linearly with time until rupture occurs. In our {\it force-ramp} experiments {\it in-silico}, the model displays the well known `mechanical signatures' of a catch-bond probed by molecular force spectroscopy. Exploiting this new evidence, we have further strengthened the analogy between MT-kt attachments and common ligand-receptor bonds in spite of the crucial differences in their underlying physical mechanisms. We then extend the formalism to model the stochastic kinetics of an attachment formed by a bundle of multiple parallel microtubules with a single kt considering the effect of rebinding under force-clamp and force-ramp conditions. From numerical studies of the model we predict the trends of variation of the mean life time and mean rupture force with the increasing number of MTs in the bundle. Both the mean life time and the mean rupture force display nontrivial nonlinear dependence on the maximum number of MTs that can attach simultaneously to the same kt.




 A new class of formal latent-variable stochastic processes called hidden quantum models (HQM's) is defined in order to clarify the theoretical foundations of ion channel signal processing. HQM's are based on quantum stochastic processes which formalize time-dependent observation. They allow the calculation of autocovariance functions which are essential for frequency-domain signal processing. HQM's based on a particular type of observation protocol called independent activated measurements are shown to to be distributionally equivalent to hidden Markov models yet without an underlying physical Markov process. Since the formal Markov processes are non-physical, the theory of activated measurement allows merging energy-based Eyring rate theories of ion channel behavior with the more common phenomenological Markov kinetic schemes to form energy-modulated quantum channels. Using the simplest quantum channel model consistent with neuronal membrane voltage-clamp experiments, activation eigenenergies are calculated for the Hodgkin-Huxley K+ and Na+ ion channels. It is also shown that maximizing entropy under constrained activation energy yields noise spectral densities approximating $S(f) \sim 1/f^\alpha$, thus offering a biophysical explanation for the ubiquitous $1/f$-type in neurological signals.
A new class of formal latent-variable stochastic processes called hidden quantum models (HQM's) is defined in order to clarify the theoretical foundations of ion channel signal processing. HQM's are based on quantum stochastic processes which formalize time-dependent observation. They allow the calculation of autocovariance functions which are essential for frequency-domain signal processing. HQM's based on a particular type of observation protocol called independent activated measurements are shown to to be distributionally equivalent to hidden Markov models yet without an underlying physical Markov process. Since the formal Markov processes are non-physical, the theory of activated measurement allows merging energy-based Eyring rate theories of ion channel behavior with the more common phenomenological Markov kinetic schemes to form energy-modulated quantum channels. Using the simplest quantum channel model consistent with neuronal membrane voltage-clamp experiments, activation eigenenergies are calculated for the Hodgkin-Huxley K+ and Na+ ion channels. It is also shown that maximizing entropy under constrained activation energy yields noise spectral densities approximating $S(f) \sim 1/f^\alpha$, thus offering a biophysical explanation for the ubiquitous $1/f$-type in neurological signals.

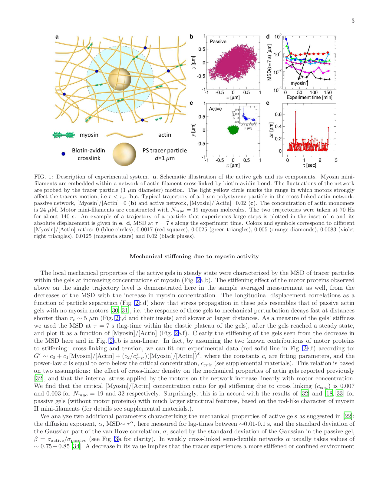
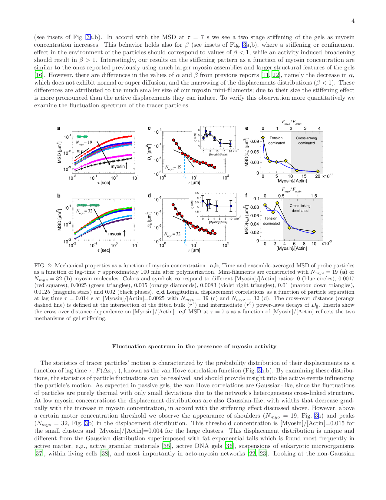
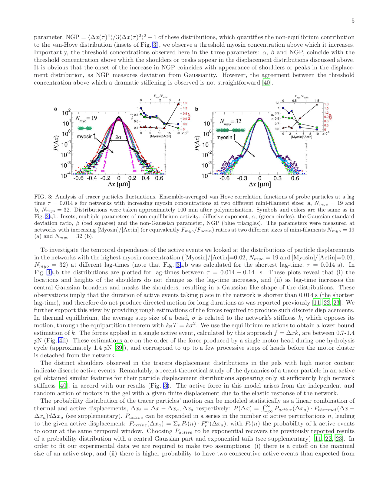
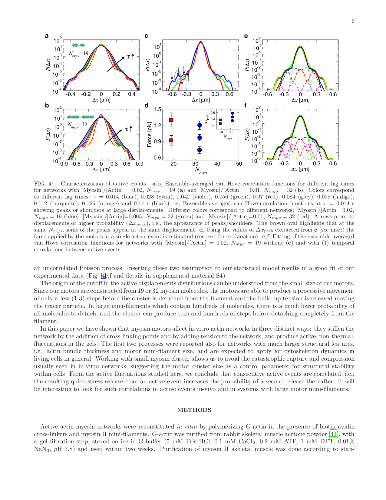 The cytoskeleton protein actin assembles into large bundles when supporting stresses in the cell, but grows into a fine branched network to induce cell motion. Such self-organization processes are studied in artificial networks of cytoskeleton proteins with thick actin bundles and large motor protein aggregates to enable optical observation. The effect of motor aggregate size on the cytoskeleton mechanical properties is studied here in networks comprised of much smaller motor assemblies. Large motor protein clusters are known to increase the stiffness of actin based networks by introducing tension and additional cross-linking cites. We find that these effects are universal to actin gels regardless of actin bundle thickness and motor aggregate size and are relevant, therefore, to a wide range of cytoskeleton based cellular processes. In contrast, motor induced active fluctuations depend significantly on motor assembly size, featuring unique non-Gaussian statistics at high concentrations of small assemblies.
The cytoskeleton protein actin assembles into large bundles when supporting stresses in the cell, but grows into a fine branched network to induce cell motion. Such self-organization processes are studied in artificial networks of cytoskeleton proteins with thick actin bundles and large motor protein aggregates to enable optical observation. The effect of motor aggregate size on the cytoskeleton mechanical properties is studied here in networks comprised of much smaller motor assemblies. Large motor protein clusters are known to increase the stiffness of actin based networks by introducing tension and additional cross-linking cites. We find that these effects are universal to actin gels regardless of actin bundle thickness and motor aggregate size and are relevant, therefore, to a wide range of cytoskeleton based cellular processes. In contrast, motor induced active fluctuations depend significantly on motor assembly size, featuring unique non-Gaussian statistics at high concentrations of small assemblies.




 We propose a stochastic model for gene transcription coupled to DNA supercoiling, where we incorporate the experimental observation that polymerases create supercoiling as they unwind the DNA helix, and that these enzymes bind more favourably to regions where the genome is unwound. Within this model, we show that when the transcriptionally induced flux of supercoiling increases, there is a sharp crossover from a regime where torsional stresses relax quickly and gene transcription is random, to one where gene expression is highly correlated and tightly regulated by supercoiling. In the latter regime, the model displays transcriptional bursts, waves of supercoiling, and up-regulation of divergent or bidirectional genes. It also predicts that topological enzymes which relax twist and writhe should provide a pathway to down-regulate transcription. This article has been published in Physical Review Letters, May 2016.
We propose a stochastic model for gene transcription coupled to DNA supercoiling, where we incorporate the experimental observation that polymerases create supercoiling as they unwind the DNA helix, and that these enzymes bind more favourably to regions where the genome is unwound. Within this model, we show that when the transcriptionally induced flux of supercoiling increases, there is a sharp crossover from a regime where torsional stresses relax quickly and gene transcription is random, to one where gene expression is highly correlated and tightly regulated by supercoiling. In the latter regime, the model displays transcriptional bursts, waves of supercoiling, and up-regulation of divergent or bidirectional genes. It also predicts that topological enzymes which relax twist and writhe should provide a pathway to down-regulate transcription. This article has been published in Physical Review Letters, May 2016.
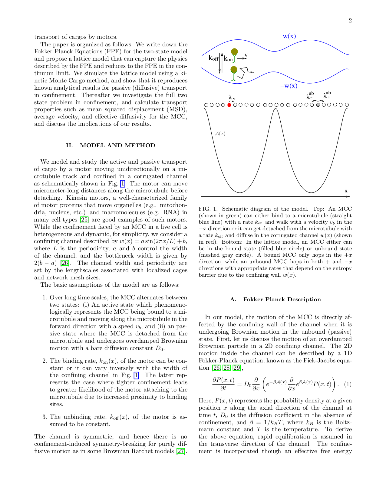

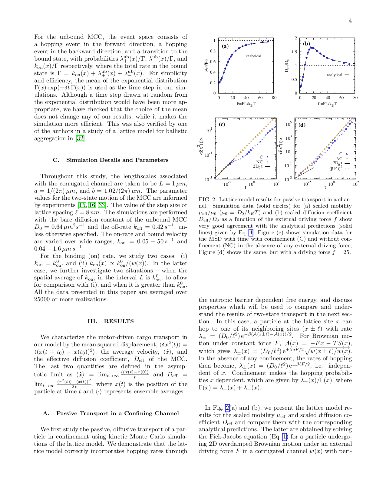
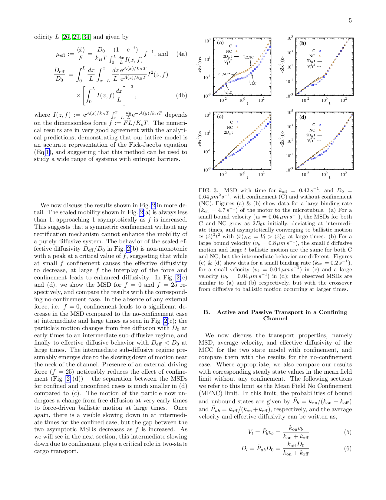
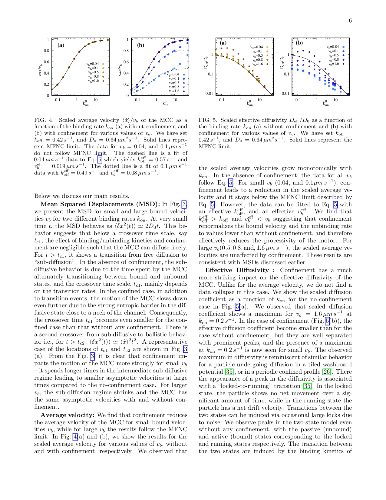 Inside cells, cargos such as vesicles and organelles are transported by molecular motors to their correct locations via active motion on cytoskeletal tracks and passive, Brownian diffusion. During the transportation of cargos, motor-cargo complexes (MCC) navigate the confining and crowded environment of the cytoskeletal network and other macromolecules. Motivated by this, we study a minimal two-state model of motor-driven cargo transport in confinement and predict transport properties that can be tested in experiments. We assume that the motion of the MCC is directly affected by the entropic barrier due to confinement if it is in the passive, unbound state, but not in the active, bound state where it moves with a constant bound velocity. We construct a lattice model based on a Fokker Planck description of the two-state system, study it using a kinetic Monte Carlo method and compare our numerical results with analytical expressions for a mean field limit. We find that the effect of confinement strongly depends on the bound velocity and the binding kinetics of the MCC. Confinement effectively reduces the effective diffusivity and average velocity, except when it results in an enhanced average binding rate and thereby leads to a larger average velocity than when unconfined.
Inside cells, cargos such as vesicles and organelles are transported by molecular motors to their correct locations via active motion on cytoskeletal tracks and passive, Brownian diffusion. During the transportation of cargos, motor-cargo complexes (MCC) navigate the confining and crowded environment of the cytoskeletal network and other macromolecules. Motivated by this, we study a minimal two-state model of motor-driven cargo transport in confinement and predict transport properties that can be tested in experiments. We assume that the motion of the MCC is directly affected by the entropic barrier due to confinement if it is in the passive, unbound state, but not in the active, bound state where it moves with a constant bound velocity. We construct a lattice model based on a Fokker Planck description of the two-state system, study it using a kinetic Monte Carlo method and compare our numerical results with analytical expressions for a mean field limit. We find that the effect of confinement strongly depends on the bound velocity and the binding kinetics of the MCC. Confinement effectively reduces the effective diffusivity and average velocity, except when it results in an enhanced average binding rate and thereby leads to a larger average velocity than when unconfined.




 During the asymmetric division of the Caenorhabditis elegans nematode zygote, the polarity cues distribution and daughter cell fates depend on the correct positioning of the mitotic spindle, which results from both centering and cortical pulling forces. Revealed by anaphase spindle rocking, these pulling forces are regulated by the force generator dynamics, which are in turn consequent of mitotic progression. We found a novel, additional, regulation of these forces by the spindle position. It controls astral microtubule availability at the cortex, on which the active force generators can pull. Importantly, this positional control relies on the polarity dependent LET-99 cortical band, which restricts or concentrates generators to a posterior crescent. We ascribed this control to the microtubule dynamics at the cortex. Indeed, in mapping the cortical contacts, we found a correlation between the centrosome-cortex distance and the microtubule contact density. In turn, it modulates pulling force generator activity. We modelled this control, predicting and experimentally validating that the posterior crescent extent controlled where the anaphase oscillations started, in addition to mitotic progression. Finally, we propose that spatially restricting force generator to a posterior crescent sets the spindle's final position, reflecting polarity through the LET-99 dependent restriction of force generators to a posterior crescent. This regulation superimposes that of force generator processivity. This novel control confers a low dependence on microtubule and active force generator exact numbers or dynamics, provided that they exceed the threshold needed for posterior displacement. Interestingly, this robustness originates in cell mechanics rather than biochemical networks.
During the asymmetric division of the Caenorhabditis elegans nematode zygote, the polarity cues distribution and daughter cell fates depend on the correct positioning of the mitotic spindle, which results from both centering and cortical pulling forces. Revealed by anaphase spindle rocking, these pulling forces are regulated by the force generator dynamics, which are in turn consequent of mitotic progression. We found a novel, additional, regulation of these forces by the spindle position. It controls astral microtubule availability at the cortex, on which the active force generators can pull. Importantly, this positional control relies on the polarity dependent LET-99 cortical band, which restricts or concentrates generators to a posterior crescent. We ascribed this control to the microtubule dynamics at the cortex. Indeed, in mapping the cortical contacts, we found a correlation between the centrosome-cortex distance and the microtubule contact density. In turn, it modulates pulling force generator activity. We modelled this control, predicting and experimentally validating that the posterior crescent extent controlled where the anaphase oscillations started, in addition to mitotic progression. Finally, we propose that spatially restricting force generator to a posterior crescent sets the spindle's final position, reflecting polarity through the LET-99 dependent restriction of force generators to a posterior crescent. This regulation superimposes that of force generator processivity. This novel control confers a low dependence on microtubule and active force generator exact numbers or dynamics, provided that they exceed the threshold needed for posterior displacement. Interestingly, this robustness originates in cell mechanics rather than biochemical networks.